
Public Health
An exemplary academic environment that prepares interprofessional learners to achieve high standards of public health practice.

Clinical and Translation Research
Designed to prepare health care professionals with the academic and research skills needed to be competitive for independent research.
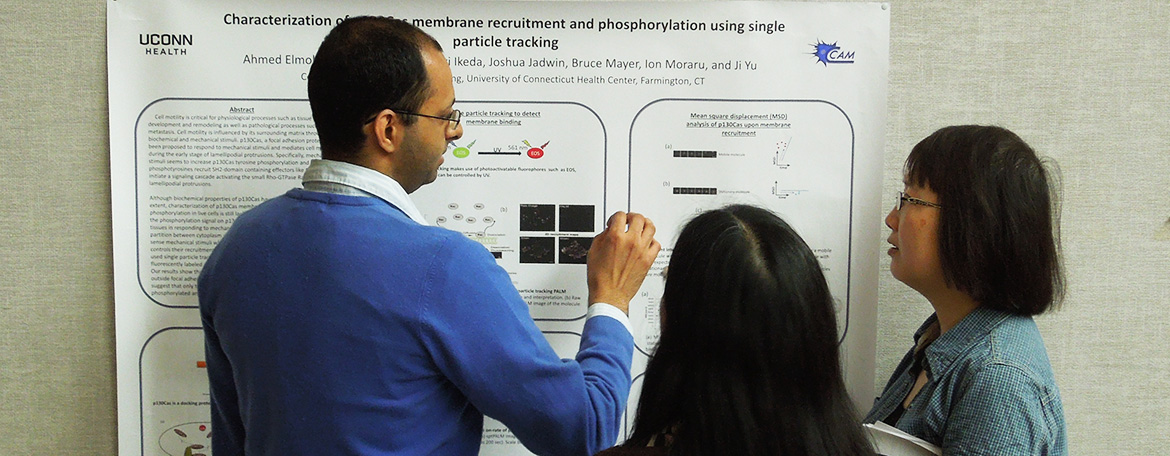
Biomedical Science
An academic environment which fosters creative thinking and supports programs leading to excellence in scholarship, research and teaching.

M.D./Ph.D.
An intellectually stimulating and highly integrated educational experience leading to completion of both the M.D. and Ph.D. degrees.

Dental Science
An interdepartmental program leading to the degree of Master of Dental Science which offers an opportunity for study and research in dental science, the basic life sciences, and the allied health fields.

D.M.D./Ph.D. Program
Designed to train individuals who are interested in pursuing research related to Oral and Craniofacial Sciences and Health and interested in careers in academic dentistry and beyond
 Dr. Renata Schiavo was the inaugural speaker at the Graduate School DEI Committee's new Seminar Series. Find out more about the series and her presentation, "The Science of Trust: Implications in Educational Settings" here.
Dr. Renata Schiavo was the inaugural speaker at the Graduate School DEI Committee's new Seminar Series. Find out more about the series and her presentation, "The Science of Trust: Implications in Educational Settings" here. Students in the Skeletal Biology Regeneration Area of Concentration in the Biomedical Science PhD program showcased their research at their Second Annual Symposium.
Students in the Skeletal Biology Regeneration Area of Concentration in the Biomedical Science PhD program showcased their research at their Second Annual Symposium. Our Ph.D. graduate program in Biomedical Science is dedicated to quality education tailored to the needs of the individual student.
Our Ph.D. graduate program in Biomedical Science is dedicated to quality education tailored to the needs of the individual student.Watch as Dr. Vanessa Scanlon, a graduate of that program now on our faculty, explains how it's contributed to her success.
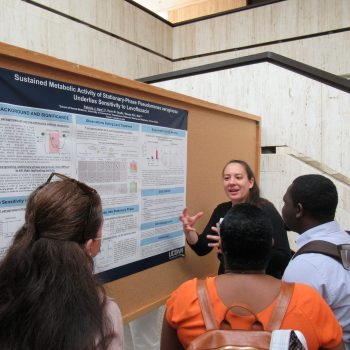 40th Annual Biomedical Science Research Day Features Program Alums
40th Annual Biomedical Science Research Day Features Program Alums Get to know 2023 UConn Health Graduate School Commencement speaker: Emily Fabrizio-Stover
Get to know 2023 UConn Health Graduate School Commencement speaker: Emily Fabrizio-Stover Read UConn President Dr. Radenka Maric's CT Mirror OpEd on the importance of diverse educational communities here.
Read UConn President Dr. Radenka Maric's CT Mirror OpEd on the importance of diverse educational communities here.
The Jackson Laboratory Collaboration

The Jackson Laboratory offers training opportunities in mammalian genetics and genomic medicine.
UConn Health Graduate School Newsletter
Learn more about UConn Health Graduate School programs in our latest newsletter: February 2024 UConn Health Graduate School Newsletter
Prospective Students: Biomedical Science PhD Program Virtual Open House
Prospective students can find more information on our program here: https://health.uconn.edu/graduate-school/prospective/
Questions about admissions? Please contact BiomedSciAdmissions@uchc.edu
About UConn Health
UConn Health, comprising the Graduate School, School of Medicine, and School of Dental Medicine, is situated on 150 acres of wooded hilltop. Our Graduate School student body consists of students from diverse backgrounds and cultures. The Graduate School at UConn Health offers programs in four fields of study – Biomedical Science, Public Health, Clinical and Translational Research, and Dental Science.
UConn Health maintains an academic environment that fosters creative thinking and provides educational opportunities tailored to the needs of individual students. Our faculty are committed to mentoring and training the next generation of scholars and scientists in a collegial and supportive environment. A variety of financial support packages are available to eligible students and affordable housing is available in several surrounding areas.
News and Announcements
UConn Health Graduate School DEI Committee Launches New Seminar Series
40th Annual Biomedical Science Graduate Student Research Day Features Program Alumni
Why don't Vaccines Work as Well in Some Older Adults?
Researchers Improve Efficacy and Efficiency of CRISPR Diagnostic Technology
UConn Participates in First Ever Clinical Trial of Ketogenic Supplements and Health Aging
Youthful Proteins Help Regrow Nerves
Science in Seconds: Handheld 3D Bioprinters to Treat Musculoskeletal Injuries
Skeletal Biology and Regeneration Students Showcase Research at Second Annual Symposium
Dr. Yulan Xiong IDs Key Mechanism in Parkinson's Disease Research
Dr. Irina Bezsonova's Research Grant Targets Hao-Foundation Syndrome
Dr. Ming Xu Receives AFAR Rising Star Award in Aging Research0
Dr. Cato T. Laurencin Named 2023 Inventor of the Year by IPOEF
Innovation at UConn Health: Meet Basic Scientist Dr. Kimberly Dodge-Kafka
Meet The Class of 2023: UConn Health Graduate School Commencement Speaker Emily Fabrizio-Stover
Science in Seconds: The Wonders of Walnuts in Colon Cancer Research
Combining Two Bone Healing Remedies Can Inhibit Bone Growth and Regeneration
Mitochondria Linked to Major Depression in Older Adults
The Heart of the Matter: New Drug Reduces Inflammation During Coronary Catheterization
Q&A: Dr. Pramod Srivastava Explains the Hunt for a Breast Cancer Vaccine